
 Anna R. C. Martin
Anna R. C. Martinanna.russocm@gmail.com
Genes identificados nas seqüências de apenas um grupo taxonômico e não em outros, tem sido naturalmente considerados como novidades evolutivas (Raible, 2004). Ou seja, alguns genes considerados exclusivos de um determinado táxon identificariam uma novidade evolutiva provavelmente responsável por originar característica nova que levará à divergência do nó ancestral, com o surgimento de novas espécies. Um exemplo seria o surgimento de simetria bilateral, cujo gene responsável provavelmente surgiu no Pré-Cambriano e levou ao aparecimento dos Bilateria.
 Figura 1 - Árvore filogenética mostrando a ramificação dos Bilateria no período Pré-Cambriano.
Figura 1 - Árvore filogenética mostrando a ramificação dos Bilateria no período Pré-Cambriano.
Após análises comparativas de genomas de algumas espécies, verificou-se que alguns genes que pareciam exclusivos de uma determinada espécie estavam presentes também em outras consideradas distantes nas hipóteses filogenéticas disponíveis. Um exemplo é Acropora (grupo de cnidário que formam corais) que se ramifica na filogenia antes de Urbilateria, porém compartilha 12% de genes exclusivos com Homo sapiens contra 1% com Drosophila melanogaster e Caenorhabditis elegans. Há duas hipóteses que explicam esse compartilhamento de genes: a primeira seria que o gene surge em Eumetazoa (presente em Acropora), porém se perde em Protostomia (representado pela ausência em C. elegans e Drosophila) mas não em Deuterostomia (presente em H. sapiens), o que nos faz concluir que H. sapiens perdeu menos genes do que as outras duas espécies de animais bilaterais e que o Urmetazoa era mais complexo do que se esperava, uma vez que esses genes já eram expressos nele. A outra hipótese é que o mesmo gene tenha aparecido no H. sapiens e em Acropora independentemente. No entanto, se considerarmos o princípio da parcimônia, o gene pode ser perdido facilmente mas é improvável que ele surja exatamente igual e independentemente em duas espécies distintas e filogeneticamente distantes (mas não impossível, claro).
 Figura 2 – Filogenia de espécies de eucariotos baseada na perda genética (modificado de Kortschak et al. 2003).
Figura 2 – Filogenia de espécies de eucariotos baseada na perda genética (modificado de Kortschak et al. 2003).
O princípio de parcimônia reforça a teoria da perda gênica, que considera que um gene compartilhado por um grupo de espécies pode desaparecer independentemente em cada espécie ou grupo. Um dos motivos para acontecer a perda gênica é quando um gene é pouco acionado. Desta forma, genes com a menor propensão para a perda gênica, durante a evolução, parecem estar mais envolvidos em funções indispensáveis do que os genes que foram perdidos em diferentes linhagens (Krylov et. al. 2004). Portanto, se um gene é mantido durante um longo período na evolução, é de se esperar que sua função seja essencial para a sobrevivência daquela espécie. Por isso que, em uma análise comparativa de genomas, o padrão de conservação de um determinado gene pode ser indicador de genes que expressam funções essenciais. Isso implica que genes expressos evoluem mais lentamente, ou seja, há mais conservação e reparo gênico, impedindo que ele sofra mutações e se perca.
Um exemplo de linhagem que provavelmente sofreu muitas perdas gênicas durante sua evolução é a que deu origem ao Trichoplax adherens, única espécie que compõe o filo Placozoa. Essa espécie pouco conhecida foi descoberta em 1883 em um aquário de água salgada no Instituto Zoológico de Graz, na Austrália. Seu corpo possui de 2 a 3 mm de diâmetro, sendo um organismo morfologicamente simples composto apenas por uma placa de camada dupla de células. Esse organismo tão simples é importante para a compreensão da evolução pelo fato de que ele possui um sistema digestório provisório no qual a camada de células ventral serve como “estômago” após sofrer uma invaginação, que engloba o alimento a ser digerido. Por ter esse sistema, os Placozoa são inseridos na filogenia de Metazoa, por alguns biólogos, entre Porifera e Cnidaria. Mesmo esse posicionamento é controverso, pelo fato do Trichoplax ser um organismo extremamente simples e pequeno, mais até que os próprios poríferos. Além disso, o Trichoplax possui um DNA pequeno (aproximado com o tamanho de uma bactéria ou um protista), o que faz pensar que muitos genes foram perdidos durante a evolução do grupo.
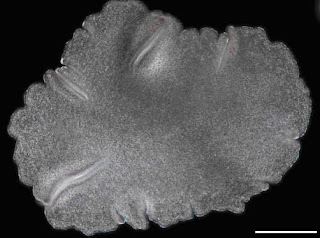 Figura 3 - Trichoplax (modificado de Srivastava et al., 2008)
Figura 3 - Trichoplax (modificado de Srivastava et al., 2008)
O estudo da perda gênica começou através de análises comparativas de conjuntos de genes de genomas completamente sequenciados – atualmente, existem poucas espécies com o DNA completamente sequenciado. Essas espécies são chamadas de organismos-modelo ou espécies-modelo e são usadas constantemente em analises comparativas de genomas. Hoje, tem-se o genoma completo da Arabidopsis tatiana (planta), C. elegans, D. melanogaster e H. sapiens (metazoários), Saccharomyces cerevisae, S. pombe (fungos) e Encephalitozoon cuniculi (protozoário), que aparecem usualmente em muitas filogenias de diferentes autores.
O problema de análises filogenéticas usando apenas organismos-modelo é que o espaço amostral é pequeno. A quantidade de espécies com genoma completamente sequenciado e sua diversidade levam a relações de parentescos que vão de encontro com relações filogenéticas já aceitas. Como exemplo, a filogenia da figura 2, que considera H. sapiens mais próximo de D. melanogaster, em relação ao C. elegans (espécie de nematóide). Isso é contrário à diversas outras propostas, que aproximam os nematóides dos insetos. Portanto, comparações feitas com base apenas nestes organismos podem dar uma imagem enganosa das relações de parentesco. Isso porque se analisarmos somente o genoma de duas espécies, pode-se encontrar seqüência semelhantes que aproximam essas duas espécies, mesmo elas sendo morfologicamente muito distintas, como a Acropora e o H. sapiens. Também pode acontecer de os genomas de espécies morfologicamente semelhantes apresentarem pouca similaridade - se a filogenia se baseasse apenas na similaridade genômica, as espécies certamente estariam afastadas filogeneticamente. Isso ocorre porque não está sendo considerado que, ao longo da evolução, pode ter ocorrido uma série de perdas gênicas e mutações que podem levar à divergência de duas sequências gênicas ou manter próximas espécies nas quais ocorreram possíveis perdas.
 Figura 4 - Árvore filogenética com todos os metazoários apontando a posição de cada espécie animal da filogenia presente na figura 2.
Figura 4 - Árvore filogenética com todos os metazoários apontando a posição de cada espécie animal da filogenia presente na figura 2.
Uma das possibilidades de diminuir conclusões errôneas sobre as relações de parentesco é analisar as sequências de genes juntamente com a função que ele exerce em cada grupo de metazoários. Análises genéticas recentes mostraram que existem genes homólogos presentes em todos metazoários que são responsáveis por seu desenvolvimento. Esses genes são conhecidos como “caixa de ferramentas” ou genes do desenvolvimento e são responsáveis por ativar outros genes que podem dar origem a apêndices, por exemplo.
Estudos mais completos utilizando dados moleculares nos permitem entender o porquê de alguns metazoários não terem certos genes, como o caso dos Placozoa. Também nos permite saber que, apesar de duas espécies possuírem genes relativamente semelhantes, elas não estão necessariamente próximas em uma árvore filogenética, como é o caso dos H. sapiens e a Acropora. Portanto, uma análise sobre a evolução utilizando apenas um conjunto de dados ou fonte de evidências pode ser menos conclusiva do que se interligarmos vários dos caminhos que levaram a grande diversidade de hoje.
Referências bibliográficas
Kortschak, R., Samuel, G., Saint, R., & Miller, D. (2003). EST Analysis of the Cnidarian Acropora millepora Reveals Extensive Gene Loss and Rapid Sequence Divergence in the Model Invertebrates Current Biology, 13 (24), 2190-2195 DOI: 10.1016/S0960-9822(03)00872-8
Krylov, D. M., Wolf Y. I., Rogozin I. B. & Koonin E. V., 2003, Gene loss, protein sequence divergence, gene dispensability, expression level, and interactivity are correlated in eukaryotic evolution, Genome Res. 13: 2229–2235.
Raible, F. & Arendt, D., 2004. Metazoan evolution: some animals are more equal than others. Current Biology 14: R106-R108.
Srivastava, M., Begovic E., Chapman J., Putnam N. H., Kawashima T., Kou A., Mitros T., Salamov A., Carpenter M. L., Signorovitch A.Y, Moreno M. A., Kamm K., Grimwoods J., Schmutz J., Shapiroi H., Grigorev I. V., Buss L.W., Schierwater B., Dellaporta S. L. & Rokhsar D. S., 2008. The Trichoplax genome and the nature of placozoans. Nature 454:955-960.
Genes identificados nas seqüências de apenas um grupo taxonômico e não em outros, tem sido naturalmente considerados como novidades evolutivas (Raible, 2004). Ou seja, alguns genes considerados exclusivos de um determinado táxon identificariam uma novidade evolutiva provavelmente responsável por originar característica nova que levará à divergência do nó ancestral, com o surgimento de novas espécies. Um exemplo seria o surgimento de simetria bilateral, cujo gene responsável provavelmente surgiu no Pré-Cambriano e levou ao aparecimento dos Bilateria.
 Figura 1 - Árvore filogenética mostrando a ramificação dos Bilateria no período Pré-Cambriano.
Figura 1 - Árvore filogenética mostrando a ramificação dos Bilateria no período Pré-Cambriano.Após análises comparativas de genomas de algumas espécies, verificou-se que alguns genes que pareciam exclusivos de uma determinada espécie estavam presentes também em outras consideradas distantes nas hipóteses filogenéticas disponíveis. Um exemplo é Acropora (grupo de cnidário que formam corais) que se ramifica na filogenia antes de Urbilateria, porém compartilha 12% de genes exclusivos com Homo sapiens contra 1% com Drosophila melanogaster e Caenorhabditis elegans. Há duas hipóteses que explicam esse compartilhamento de genes: a primeira seria que o gene surge em Eumetazoa (presente em Acropora), porém se perde em Protostomia (representado pela ausência em C. elegans e Drosophila) mas não em Deuterostomia (presente em H. sapiens), o que nos faz concluir que H. sapiens perdeu menos genes do que as outras duas espécies de animais bilaterais e que o Urmetazoa era mais complexo do que se esperava, uma vez que esses genes já eram expressos nele. A outra hipótese é que o mesmo gene tenha aparecido no H. sapiens e em Acropora independentemente. No entanto, se considerarmos o princípio da parcimônia, o gene pode ser perdido facilmente mas é improvável que ele surja exatamente igual e independentemente em duas espécies distintas e filogeneticamente distantes (mas não impossível, claro).
 Figura 2 – Filogenia de espécies de eucariotos baseada na perda genética (modificado de Kortschak et al. 2003).
Figura 2 – Filogenia de espécies de eucariotos baseada na perda genética (modificado de Kortschak et al. 2003).O princípio de parcimônia reforça a teoria da perda gênica, que considera que um gene compartilhado por um grupo de espécies pode desaparecer independentemente em cada espécie ou grupo. Um dos motivos para acontecer a perda gênica é quando um gene é pouco acionado. Desta forma, genes com a menor propensão para a perda gênica, durante a evolução, parecem estar mais envolvidos em funções indispensáveis do que os genes que foram perdidos em diferentes linhagens (Krylov et. al. 2004). Portanto, se um gene é mantido durante um longo período na evolução, é de se esperar que sua função seja essencial para a sobrevivência daquela espécie. Por isso que, em uma análise comparativa de genomas, o padrão de conservação de um determinado gene pode ser indicador de genes que expressam funções essenciais. Isso implica que genes expressos evoluem mais lentamente, ou seja, há mais conservação e reparo gênico, impedindo que ele sofra mutações e se perca.
Um exemplo de linhagem que provavelmente sofreu muitas perdas gênicas durante sua evolução é a que deu origem ao Trichoplax adherens, única espécie que compõe o filo Placozoa. Essa espécie pouco conhecida foi descoberta em 1883 em um aquário de água salgada no Instituto Zoológico de Graz, na Austrália. Seu corpo possui de 2 a 3 mm de diâmetro, sendo um organismo morfologicamente simples composto apenas por uma placa de camada dupla de células. Esse organismo tão simples é importante para a compreensão da evolução pelo fato de que ele possui um sistema digestório provisório no qual a camada de células ventral serve como “estômago” após sofrer uma invaginação, que engloba o alimento a ser digerido. Por ter esse sistema, os Placozoa são inseridos na filogenia de Metazoa, por alguns biólogos, entre Porifera e Cnidaria. Mesmo esse posicionamento é controverso, pelo fato do Trichoplax ser um organismo extremamente simples e pequeno, mais até que os próprios poríferos. Além disso, o Trichoplax possui um DNA pequeno (aproximado com o tamanho de uma bactéria ou um protista), o que faz pensar que muitos genes foram perdidos durante a evolução do grupo.
 Figura 3 - Trichoplax (modificado de Srivastava et al., 2008)
Figura 3 - Trichoplax (modificado de Srivastava et al., 2008)O estudo da perda gênica começou através de análises comparativas de conjuntos de genes de genomas completamente sequenciados – atualmente, existem poucas espécies com o DNA completamente sequenciado. Essas espécies são chamadas de organismos-modelo ou espécies-modelo e são usadas constantemente em analises comparativas de genomas. Hoje, tem-se o genoma completo da Arabidopsis tatiana (planta), C. elegans, D. melanogaster e H. sapiens (metazoários), Saccharomyces cerevisae, S. pombe (fungos) e Encephalitozoon cuniculi (protozoário), que aparecem usualmente em muitas filogenias de diferentes autores.
O problema de análises filogenéticas usando apenas organismos-modelo é que o espaço amostral é pequeno. A quantidade de espécies com genoma completamente sequenciado e sua diversidade levam a relações de parentescos que vão de encontro com relações filogenéticas já aceitas. Como exemplo, a filogenia da figura 2, que considera H. sapiens mais próximo de D. melanogaster, em relação ao C. elegans (espécie de nematóide). Isso é contrário à diversas outras propostas, que aproximam os nematóides dos insetos. Portanto, comparações feitas com base apenas nestes organismos podem dar uma imagem enganosa das relações de parentesco. Isso porque se analisarmos somente o genoma de duas espécies, pode-se encontrar seqüência semelhantes que aproximam essas duas espécies, mesmo elas sendo morfologicamente muito distintas, como a Acropora e o H. sapiens. Também pode acontecer de os genomas de espécies morfologicamente semelhantes apresentarem pouca similaridade - se a filogenia se baseasse apenas na similaridade genômica, as espécies certamente estariam afastadas filogeneticamente. Isso ocorre porque não está sendo considerado que, ao longo da evolução, pode ter ocorrido uma série de perdas gênicas e mutações que podem levar à divergência de duas sequências gênicas ou manter próximas espécies nas quais ocorreram possíveis perdas.
 Figura 4 - Árvore filogenética com todos os metazoários apontando a posição de cada espécie animal da filogenia presente na figura 2.
Figura 4 - Árvore filogenética com todos os metazoários apontando a posição de cada espécie animal da filogenia presente na figura 2.Uma das possibilidades de diminuir conclusões errôneas sobre as relações de parentesco é analisar as sequências de genes juntamente com a função que ele exerce em cada grupo de metazoários. Análises genéticas recentes mostraram que existem genes homólogos presentes em todos metazoários que são responsáveis por seu desenvolvimento. Esses genes são conhecidos como “caixa de ferramentas” ou genes do desenvolvimento e são responsáveis por ativar outros genes que podem dar origem a apêndices, por exemplo.
Estudos mais completos utilizando dados moleculares nos permitem entender o porquê de alguns metazoários não terem certos genes, como o caso dos Placozoa. Também nos permite saber que, apesar de duas espécies possuírem genes relativamente semelhantes, elas não estão necessariamente próximas em uma árvore filogenética, como é o caso dos H. sapiens e a Acropora. Portanto, uma análise sobre a evolução utilizando apenas um conjunto de dados ou fonte de evidências pode ser menos conclusiva do que se interligarmos vários dos caminhos que levaram a grande diversidade de hoje.
Referências bibliográficas
Kortschak, R., Samuel, G., Saint, R., & Miller, D. (2003). EST Analysis of the Cnidarian Acropora millepora Reveals Extensive Gene Loss and Rapid Sequence Divergence in the Model Invertebrates Current Biology, 13 (24), 2190-2195 DOI: 10.1016/S0960-9822(03)00872-8
Krylov, D. M., Wolf Y. I., Rogozin I. B. & Koonin E. V., 2003, Gene loss, protein sequence divergence, gene dispensability, expression level, and interactivity are correlated in eukaryotic evolution, Genome Res. 13: 2229–2235.
Raible, F. & Arendt, D., 2004. Metazoan evolution: some animals are more equal than others. Current Biology 14: R106-R108.
Srivastava, M., Begovic E., Chapman J., Putnam N. H., Kawashima T., Kou A., Mitros T., Salamov A., Carpenter M. L., Signorovitch A.Y, Moreno M. A., Kamm K., Grimwoods J., Schmutz J., Shapiroi H., Grigorev I. V., Buss L.W., Schierwater B., Dellaporta S. L. & Rokhsar D. S., 2008. The Trichoplax genome and the nature of placozoans. Nature 454:955-960.


Olá Anna!
ResponderExcluirMuito interessante seu texto e você levanta alguns pontos muito interessantes: homologia, caracteres (bases de dados) e filogenias.
O problema das homologias não é recente em sistemática. Algumas proposições, como homologias primárias e secundárias (p.ex. de Pinna, 1991), tornaram-se ícones e acabam dificultando a inserção de novos pensamentos em um panorama evolutivo. Hoje, com o desenvolvimento da biologia do desenvolvimento e da evo-devo, é possível pensar em homologias profundas, em características homoplásticas como resultado de um processo evolutivo do qual as características são sim compartilhadas em um nível mais remoto da filogenia, e não mais como erro procedimental. Os genes Hox (referidos por você como "caixa de ferramentas") mostram que o padrões corporais até então exclusivos de linhagens filogenéticas distantes (como artrópodes e vertebrados) são na verdade determinados por um conjunto de genes homólogos e compartilhados pelos mais variados níveis da filogenia. Isso mostra que quanto mais variada for a base de dados utilizada, melhor suportadas serão as hipóteses de filogenia resultantes, mas também levanta a questão sobre a realização de análises conjuntas (bases de dados diferentes analisadas simultaneamente) ou individuais, com comparações posteriores entre as topologias obtidas. Nesse sentido eu concordo com você, a comparação entre filogenias resultantes de bases de dados diferentes talvez seja a única maneira de compreendermos a enorme diversidade atual, o que reforça a sistemática com área unificadora da biologia e a base da biologia comparada.
Parabéns!
Abraços!
muito legal seu comentario principalmente o que diz respeito ao placozoario que e pouco conhecido ,e muito simples .voçe sabia que ele estar na lista dos animais mais simples do mumdo legaaaaaaaaaaaaaa
ResponderExcluir